Les coquilles des escargots sont des spirales qui peuvent croître de manière dextre ou senestre. En fait, ils sont presque tous dextres. Seuls un sur dix mille est senestre dans l’espèce des petits gris mais il existe des espèces où c’est le contraire.
Un petit gris. Si on place sa tête à gauche, sa coquille s’enroule dans le sens trigonométrique.
Les tire-bouchons usuels, c’est-à-dire pour droitiers, sont dextres, les tire-bouchons pour gauchers sont senestres.
Un tire-bouchon dextre.
Les queues de cochons
De même la queue en tire-bouchon des cochons peut être dextre ou senestre. Dans ce cas, il se trouve qu’il y a autant de cochons dextres que de cochons senestres. Le sexe des cochons mâles a la même propriété. Ces différences entre dextre et senestre se retrouvent au niveau des molécules, ce qui a parfois des conséquences sur leurs propriétés.
Le pangolin en boule
Pour échapper à ses prédateurs, le pangolin (oui, le mammifère impliqué dans l’origine du Covid-19) se roule en boule, ce qui le rend vulnérable aux braconniers.
Pangolin en boule
D’où la question : existe-t-il deux sortes de pangolins selon l’orientation de la boule ?
Jean-Henri Fabre est connu pour son observation des insectes. Excellent vulgarisateur, il est de ceux qui savent communiquer leurs passions. Les mathématiques en font partie.
Jean-Henri Fabre
Jean-Henri Fabre (1823 – 1915)
Bien que titulaire d’une licence de mathématiques, d’un doctorat en sciences naturelles et de plusieurs autres diplômes, Jean-Henri Fabre est un autodidacte comme il le rappelle lui-même :
Apprendre sous la direction d’un maître m’a été refusé. J’aurais tort de m’en plaindre. L’étude solitaire a sa valeur ; elle ne vous coule pas dans un moule officiel, elle vous laisse votre pleine originalité. Le fruit sauvage, s’il arrive à maturité, a une autre saveur que le produit de serre chaude ; il laisse aux lèvres qui savent l’apprécier un mélange d’amertume et de douceur dont le mérite s’accroît par le contraste.
Son côté autodidacte le rend attachant pour certains et agaçant pour d’autres. Quelques modernes lui reprochent aussi de ne pas avoir épousé les thèses de Darwin qui, en revanche, reconnaissait en lui un observateur incomparable. Il s’explique lui-même dans une de ses lettres à Darwin :
Vous vous étonnez de mon peu de goût pour les théories, si séduisantes qu’elles soient. Ce travers d’esprit, si c’en est un, tient un peu à mes longues études mathématiques qui m’ont habitué à ne reconnaître la vérité qu’à la lueur d’un irrésistible faisceau de lumière. Ne jurant par aucun maître, libre d’idées préconçues, peu enclin aux séductions des théories, je cherche avec passion la vérité, près à l’admettre quelle qu’elle soit et de quelque fait qu’elle vienne. Et comme moyen de recherche, je ne connais qu’une chose : l’expérience.
Par ailleurs, Darwin l’avait chargé d’expériences sur les insectes retournant à leurs nids. Les résultats se trouvent dans l’œuvre de Fabre. De façon générale, on trouvera la plupart des écrits de Fabre sur internet.
Fabre créationniste ?
Parmi les critiques modernes faites à Jean-Henri Fabre, certains le stigmatisent comme créationniste car il ne croyait pas à la théorie de Darwin, qu’il comparait à celle de la génération spontanée. À la défense de Fabre, il faut noter que la théorie originelle de Darwin n’était pas celle qui porte son nom aujourd’hui. Il s’agissait plutôt d’une transposition de la sélection des espèces domestiques, pratiquée depuis longtemps par les éleveurs, en une sélection naturelle sous l’effet de modifications du milieu. Autrement dit, il lui manquait l’explication qui viendra avec la découverte des gênes, par Gregor Mendel au début du XXe siècle. La théorie de l’évolution telle que nous la connaissons est postérieure de vingt ans à la mort de Fabre ! Comment peut-on lui reprocher de ne pas l’avoir reconnue ?
Mais l’essentiel n’est pas là, il est dans deux choses, sans parler de l’inélégance d’attaquer les morts, qui ne peuvent se défendre. Premièrement, il faut savoir ne pas se tromper d’adversaires. Les obscurantistes que sont les créationnistes ne sont pas les disciples de Jean-Henri Fabre. Ils sont dans des religions qui refusent la science, et malheureusement pas la violence. Deuxièmement, de Jean-Henri Fabre retenons plutôt l’exceptionnel talent de vulgarisateur. Pour finir sur une note poétique et liée à la question de l’évolution, voici l’un de ses commentaires sur la parade nuptiale des scorpions languedociens : La colombe a, dit-on, inventé le baiser. Je lui connais un précurseur : c’est le scorpion.
Parade amoureuse de scorpions languedociens. Dans un cas sur deux au moins, le mâle (à droite) finira dévoré par la femelle (à gauche), ce qui atténue l’impression romantique donnée par Fabre.
Jean-Henri Fabre a réussi à me faire regarder les scorpions autrement, c’est pourquoi je me souviens de cette remarque. Aujourd’hui, elle me fait m’interroger : selon la théorie créationniste, parler de précurseur d’une espèce a-t-il un sens ?
Descriptions et mathématiques chez Fabre
Dans ses souvenirs entomologiques, Jean-Henri Fabre dépeint les mœurs des insectes de manière vivante, en les ramenant souvent aux nôtres. Il décrit ainsi le carabe doré en nous emmenant d’abord visiter les abattoirs de Chicago pour comparer ensuite leur efficacité à celles des carabes dont on saisit mieux ainsi la férocité comme la voracité.
Le carabe doré, qui sera l’occasion d’une digression sur les mœurs humaines pour Jean-Henri Fabre.
Il conclut alors sur nos origines et notre avenir, avec l’abolition de l’esclavage et l’instruction des femmes, les deux voies du progrès moral selon lui. Cette façon de généraliser sera parfois critiquée plus tard, comme peu scientifique. Il est vrai que, par moments, Fabre concluait un peu vite. Par exemple, voici comment il décrit la toile d’une araignée, l’épeire :
Nous reconnaîtrons d’abord que les rayons sont équidistants ; ils forment de l’un à l’autre des angles sensiblement égaux […] les divers tours de spire […] avec les deux rayons qui les limitent, forment d’un côté un angle obtus et de l’autre un angle aigu […] d’un secteur à l’autre, ces mêmes angles, l’obtus comme l’aigu, ne changent pas de valeur, autant que peuvent en juger les scrupules du regard seul.
Fabre reconnaît alors une propriété caractéristique de la spirale logarithmique et en conclut que la toile de l’épeire épouse cette forme, ce qui est rapide surtout quand la mesure a été faite à l’œil. Ceci dit, cela n’enlève rien à la qualité de son travail, et il n’en reste pas moins que, du fait de sa construction, la toile prend une forme de spirale.
Scarabée sacré en train de confectionner une boule.
De même, c’est de manière très mathématique qu’il explique la forme de poire que le scarabée sacré donne à la bouse dans laquelle il dépose son œuf : une sphère pour minimiser la surface externe afin de réduire la dessiccation, qui rendrait la bouse immangeable pour la larve, coiffée d’une sorte de cylindre contenant l’œuf, qui se trouve ainsi dans un endroit plus aéré.
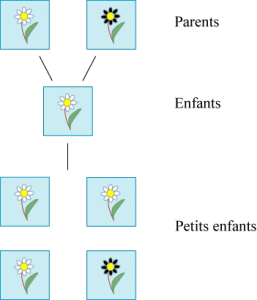
Gregor Mendel (1822 – 1884) est connu pour avoir posé les premières lois de la génétique. Elles sont de nature si mathématique que Godfrey Hardy, le grand mathématicien britannique du début XXe siècle, connu pour sa critique des mathématiques appliquées, les a prolongées. Imaginons qu’une fleur vienne en deux couleurs : blanche et noire, jamais grise ou autre et que ces deux variétés puissent s’hybrider, c’est-à-dire se mélanger. Imaginons que deux parents à fleurs blanches donnent toujours des enfants à fleurs blanches, alors que les parents à fleurs noires peuvent donner des blanches comme des noires.
Né Johann Mendel en 1822, Mendel prendra le prénom de Gregor à son entrée au monastère de Brunn (Tchéquie), en Autriche à l’époque. Il y trouva un milieu intellectuel stimulant et put y installer un jardin expérimental, où il fit ses recherches sur l’hybridation. En 1866, il devint supérieur de son couvent, ce qui mit fin à ses recherches en botanique. Il se consacra alors à l’administration du monastère ainsi qu’à des recherches en météorologie, pour lesquelles il fut reconnu par ses contemporains … davantage que pour ses apports à la génétique.
La mathématique de l’hybridation
Gregor Mendel a étudié ces lois de l’hybridation en pollinisant artificiellement des pois, qui se présentent sous deux formes facilement discernables. Nous ne décrirons pas ses expériences en détail. Son premier résultat est d’ordre statistique. En croisant une fleur noire et une fleur blanche, à la première génération, on obtient des fleurs blanches et, à la seconde, trois quarts de fleurs blanches et un quart de fleurs noires.
Deux premières générations d’un croisement blanche / noire.
Pour le mathématicien, une explication logique est de penser que le gène de la couleur des fleurs se divise en deux moitiés, ses deux allèles : blanc et noir. A priori, il existe donc quatre combinaisons possibles de ces deux allèles : blanc / blanc, blanc / noir, noir / blanc et noir / noir. Cette propriété est cachée car seuls les porteurs du gène blanc / blanc ont des fleurs blanches, tous les autres ont des fleurs noires. C’est pourquoi on parle de caractère dominant pour la couleur noire, et de caractère récessif pour la couleur blanche. Cette domination est cependant très relative car les combinaisons se faisant de façon équiprobable, à la seconde génération, nous trouvons une fois sur quatre la combinaison blanc / blanc, donc des fleurs blanches.
Cette théorie de Mendel ne fut pas comprise en son temps. Les biologistes pensaient que les caractères dominants devaient forcément augmenter dans la population, ce que les calculs précédents nient. Plus étrangement encore, on ne vit pas immédiatement le lien avec la théorie de l’évolution de Darwin, pourtant contemporaine de celle de Mendel.
Le principe de Hardy
À l’opposé de Mendel, qui était prêtre, Godfrey Hardy était un athée convaincu. Son athéisme comprenait cependant une étrange part d’autodérision, si on en croît l’anecdote suivante. La peur d’un naufrage lui fit écrire à un collègue pour lui annoncer qu’il avait démontré l’hypothèse de Riemann. Il aurait ensuite justifié son envoi en disant que Dieu, qu’il tenait pour son ennemi intime, n’allait pas le laisser mourir et laisser croire ainsi qu’un tel impie avait réussi à démontrer cette conjecture, encore ouverte de nos jours. Il est tout aussi étrange qu’un mathématicien pur aussi convaincu ait publié un article de biologie. Il le serait encore davantage si, un jour, il était plus connu pour son apport à la génétique que pour ses théorèmes mathématiques. Le moteur de recherche Google laisse penser que ce jour viendra puisque « théorème Hardy » donne 56 700 résultats alors que « principe Hardy » en donne 4 450 000. Godfrey Hardy y verrait sans doute une revanche de son ennemi.
Si les mathématiques appliquées ont pu un jour être vues comme « impures » par certains mathématiciens « purs », ce fut le cas de Godfrey Hardy. On s’étonnera alors de voir son nom mêlé à une question de biologie. C’est pourquoi il s’excusa presque de s’immiscer dans ce domaine. En 1908, au cours d’un dîner, on lui demanda s’il était possible de déterminer mathématiquement la proportion d’allèles dominants permettant l’évolution dans une population. Hardy étant un mathématicien pur, sa réponse réclama quelques hypothèses. Tout d’abord, la population devait être de grande taille, sans migration, estimée infinie, les individus s’y croiseraient aléatoirement mais les générations seraient séparées. Enfin, il n’y aurait ni mutation, ni sélection. Tout ceci assure la rigueur du raisonnement suivant.
Considérons un gène à deux allèles A et a possédant les fréquences p et q = 1 – p dans une certaine génération. Quelles sont les fréquences à la génération suivante ?
Pour le déterminer, comptons d’abord les fréquences des diverses combinaisons à la génération suivante : AA, Aa et aa. Il s’agit d’une question élémentaire de probabilité. Pour qu’un individu soit AA, il doit avoir reçu l’allèle A de ses deux parents, supposés aléatoires d’après l’hypothèse de Hardy. La fréquence de chacun étant égale à p, la probabilité est égale à p2. De même, celle de aa est q2. Pour Aa, deux cas sont possibles puisque cela peut provenir d’un A de la mère et d’un a du père, comme du contraire. On obtient donc 2 pq.
Si la population totale de cette nouvelle génération est égale à N, le nombre d’allèles y est égal à 2N. L’allèle A se trouve deux fois dans AA et une fois dans Aa, son nombre est donc égal à 2 p2 N + 2 pq N. Sa fréquence est ainsi égale à p2 + pq = p (p + q) = p puisque p + q = 1. Il en est de même de l’allèle a. Autrement dit, sous les hypothèses énoncées plus haut, la fréquence des allèles ne se modifie pas d’une génération à l’autre.
Ainsi, les relations de dominance entre allèles n’influent pas sur leurs fréquences. Autrement dit, l’évolution est impossible sous les hypothèses de Hardy … il faut tenir compte des mutations.
La même courbe se retrouve-t-elle dans les galaxies, certains mollusques et les toiles d’araignées ? Enquête sur la spirale logarithmique.
La spirale d’Archimède
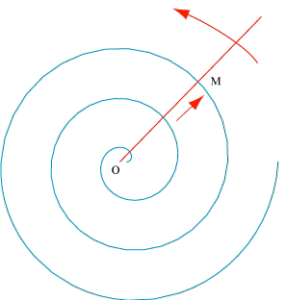
Imaginez ! Une droite tourne à vitesse angulaire constante autour d’un point O. Si, partant de O, un point M parcourt cette droite à vitesse constante, on obtient une spirale d’Archimède. On démontre facilement que les spires y sont régulièrement espacées.
Spirale d’Archimède. Elle est engendrée par un point mobile M partant d’un point O, à vitesse constante sur une droite tournant à vitesse angulaire constante autour de O.
La spirale logarithmique
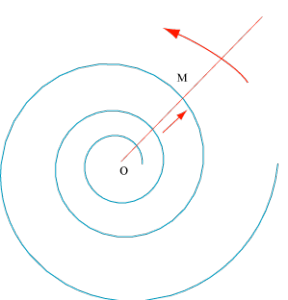
Si, toujours partant de O, le point M parcourt la droite à une vitesse proportionnelle à la longueur OM, il dessine une autre courbe, appelée spirale logarithmique depuis Pierre Varignon (1654 – 1722) mais étudiée auparavant par René Descartes (1596 – 1650) avant d’être choisie par Jacques Bernoulli (1654 – 1705) pour orner sa tombe. Malheureusement, le sculpteur ignorait cette courbe et grava une spirale d’Archimède.
Spirale logarithmique. Elle est engendrée par un point mobile M partant d’un point O, à vitesse proportionnelle à OM sur une droite tournant à vitesse angulaire constante autour de O.
Au lieu d’être régulièrement espacées, les spires suivent une progression géométrique de raison constante. Autre propriété de la spirale : elle coupe le rayon OM suivant un angle constant.
Inscription sur la tombe de Jacques Bernoulli, avec la spirale en bas.Sur cet agrandissement, on voit que le sculpteur a gravé une spirale d’Archimède et non une spirale logarithmique. L’inscription latine “eadem mutata resurgo” signifie “déplacée, je réapparais à l’identique”.
Le développement du nautile
Le nautile est un mollusque marin dont la coquille est en forme de spirale. L’espace entre les spires étant triplé à chaque enroulement, elle évoque une spirale logarithmique. Pour examiner si cette forme est fortuite ou non, il est nécessaire d’en comprendre la provenance.
Coupe d’un nautile faisant apparaître une forme de spirale logarithmique.
La coquille du nautile est divisée en chambres closes, l’animal n’occupant que la dernière. Les autres sont remplies d’un mélange de liquide et de gaz, toutes communiquent entre elles au moyen d’un siphon.
Nautile vivant. L’animal n’occupe que la dernière chambre. Il se déplace d’avant en arrière en expulsant de l’eau du côté de sa bouche.
Ces chambres correspondent à l’évolution progressive du mollusque. Quand il grossit, ne pouvant agrandir la chambre où il se trouve, il en crée une autre dans son prolongement, un peu plus grosse mais semblable.
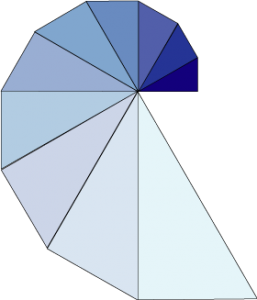
Pour montrer que cette idée mène effectivement à une spirale logarithmique, prenons comme modèle de la coquille une suite de triangles rectangles d’angle au sommet constant égal à 30°. Le rapport entre un triangle et son suivant est de 115 % (l’inverse du cosinus de 30° soit 2 divisé par racine de 3 pour être précis), ce qui correspond bien à une spirale logarithmique. L’idée correspond à un accroissement progressif de la taille de l’animal. Il n’est pas besoin d’imaginer de plans compliqués inscrits dans les gènes du nautile pour cela, juste une façon de croître.
Suite de triangles rectangles formant une (approximation de) spirale logarithmique.
La spirale logarithmique se retrouve pour les mêmes raisons dans d’autres animaux, comme la planorbe, un escargot marin très utilisé dans les aquariums car il se nourrit d’algues et de plantes à la limite du pourrissement.
Une coquille de planorbe en forme de spirale logarithmique.
Les toiles d’araignées
La toile d’araignée est avant tout un piège destiné à attraper des insectes. Certaines espèces tissent des toiles où il est bien difficile de reconnaître la moindre régularité.
Il n’est pas facile de reconnaître la moindre courbe mathématique dans cette toile d’araignée. En revanche, sans le soleil en contre jour, il est difficile de la détecter.
Les espèces les plus communes en France, les épeires, fabriquent cependant des toiles en forme de spirales. Après avoir bâti un cadre entre quelques branches, l’araignée tisse un réseau régulier de segments rectilignes partant tous d’un même point. Un fois ce travail fini, elle forme une spirale en les reliant. Le célèbre entomologiste Jean-Henri Fabre (1823 – 1915) a voulu y reconnaître une spirale logarithmique, tout en remarquant que l’action de la pesanteur transformait chaque segment en chaînette, la forme que prend naturellement un fil pesant comme les câbles électriques ou les chaînes que l’on porte autour du cou.
Cette toile d’épeire laisse plus penser à une spirale d’Archimède qu’à une spirale logarithmique. On y remarque également les segments transformés en chaînette sous l’effet de la pesanteur.
Certaines espèces de coucous font couver leurs œufs par des oiseaux de tailles très différentes. Chacune a sa stratégie de parasitage. Certains, dont le coucou gris, semblent adapter la taille de leurs œufs à celle de ceux de leur hôte involontaire.
Le coucou gris
Le coucou gris, qui a la taille d’un pigeon, parasite des passereaux. À première vue, le scénario est simple. La femelle coucou pond un œuf dans le nid de rousserolles, de roitelets, de fauvettes, ou d’autres. Son œuf est le premier à éclore. Le petit coucou expulse alors la couvée entière du nid. Les passereaux nourrissent ensuite l’intrus jusqu’à ce qu’il soit adulte.
Jeune coucou se faisant nourrir par une rousserolle.
Stratégies du coucou
Pour atteindre son but, le couple de coucous repère puis guette un nid de passereau, de l’espèce qui les a élevés de préférence. Ce choix n’est pas toujours possible, et une erreur peut être fatale au jeune coucou. Par exemple, si la femelle pond dans un nid de granivores, son petit mourra de faim, car le coucou est insectivore.
Quand la femelle passereau a pondu, celle du coucou profite de son absence, pour gober un œuf et le remplacer rapidement par l’un des siens. Son œuf éclot avant ceux des passereaux car il a commencé à incuber dans son corps. Sitôt né, encore aveugle, le petit coucou expulse tous les œufs du nid afin d’être nourri seul par ses parents adoptifs.
Oisillon coucou jetant un œuf hors du nid.
Les mathématiques du coucou
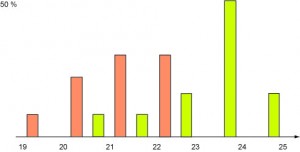
Bien que le coucou soit cinq à six fois plus grand que les passereaux qu’il parasite, sa femelle pond des œufs de taille comparable aux leurs. Plus étrange, elle semble adapter la taille de ses œufs à celle de ceux qui se trouvent dans le nid dans lequel elle pond. L’un des premiers scientifiques à avoir étudié la question quantitativement, Oswald Latter en 1902, a récolté 29 œufs de coucous dans des nids de roitelets et de fauvettes et notés les diamètres. En réunissant ces données dans deux histogrammes, nous obtenons deux courbes en cloche distinctes ce qui indique que nous avons affaire à deux populations distinctes. Autrement dit, la femelle coucou adapte bien la taille de ses œufs à ceux déjà présents dans le nid dans lequel elle pond.
Distributions des diamètres des œufs pondus dans les nids de roitelets (en orange) et de fauvettes (en vert).Cette étude a depuis été confirmée pour plusieurs espèces de coucous. La recherche est d’autant plus active sur la question que, suivant les espèces, les coucous pratiquent le parasitisme de couvée, ou non et, parmi les coucous parasites, certains sont éjecteurs (ils détruisent les œufs de leur hôte dès leur éclosion) et d’autres, non. Les seconds parasitent des espèces de taille comparable à la leur alors que les autres choisissent des oiseaux plus petits.
Le parasitisme de couvée ne se limite pas à certaines espèces de coucous. Cependant, le phénomène d’adaptation de la taille de l’œuf à celle de ceux de l’hôte n’a pas forcément lieu. Par exemple, certaines canes colverts pondent dans des nids d’autres colverts. Les flamands roses font de même ainsi que bien d’autres espèces d’oiseaux (on en a dénombré 236). Dans d’autres cas, les oiseaux parasitent des espèces de taille similaire à la leur. Seuls ceux qui parasitent des oiseaux plus petits connaissent ce phénomène d’adaptation de la taille de leurs œufs.
Les étourneaux, et d’autres oiseaux se comportent souvent comme une unité filant parfois dans une direction précise pour s’en détourner soudain. Les mouvements des bancs de poisson sont similaires. D’où viennent ces comportements ?
Un vol d’étourneaux
La défense contre les prédateurs
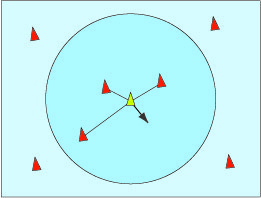
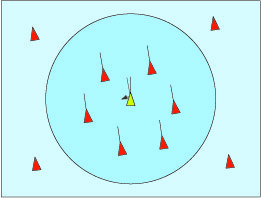
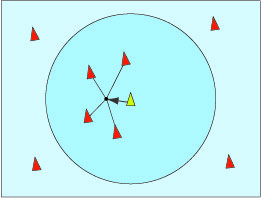
La raison essentielle de ces regroupements est la défense contre les prédateurs. Par exemple, quand les étourneaux sont effrayés, ils s’élèvent, se rassemblent et volent en formant la masse la plus compacte possible. Un rapace évite de fondre sur ce groupe de crainte de se blesser. Il cherche plutôt à sélectionner des retardataires ou des oiseaux affaiblis.
La nuée vire et tourne de telle sorte qu’il est difficile de prévoir ses mouvements, qui semblent aléatoires. De nos jours, les zoologistes sont persuadés que ce ballet ne doit rien à la présence d’un mystérieux chef d’orchestre ou à un esprit surnaturel du groupe. Dans les années 1980, Wayne Potts, professeur à l’université d’Utah, a filmé des nuées de bécasseaux pour s’apercevoir que n’importe quel individu pouvait initier un mouvement du groupe, qui se propageait ensuite très rapidement par ondes rayonnant autour de l’initiateur, et cela dans tous les sens. De plus, ces ondes se propagent bien plus rapidement que la vitesse de réaction normale d’un individu isolé peut le laisser penser. En revanche, les mouvements des oiseaux séparés du groupe ne l’influencent pas. Ils sont les cibles privilégiées des prédateurs, donc ne sont pas suivis. Cette règle a l’avantage d’accélérer la réponse du groupe à une attaque.
Un modèle mathématique
D’après l’étude de Wayne Potts, chaque oiseau réagit à ce qui l’entoure, et uniquement à cela. Son comportement peut donc être modélisé : chacun ne réagit qu’à ses voisins. En 1986, un informaticien, Craig Reynolds, précisa des règles qui simulent le comportement des nuées d’oiseaux comme celui des bancs de poissons. Il a nommé « boids » ces oiseaux virtuels (un mot à faible distance linguistique de « birds »). On peut trouver des animations sur internet utilisant son modèle (chercher Boids avec votre moteur de recherche préféré). Les trois règles sont toutes de nature locale, chaque oiseau ne réagit qu’aux mouvements de ses voisins.
Séparation
Si un oiseau est trop proche de ses voisins, il s’en écarte pour éviter les collisions.
Alignement
Alignement dans la direction du vol des oiseaux qui l’entourent.
Cohésion
Cohésion pour aller vers la position moyenne des oiseaux qui l’entourent.
Si vous voulez programmer une simulation de vol d’étourneaux, il vous reste à définir plusieurs paramètres : rayon du cercle de voisinage (en gris clair sur les figures), vitesses, accélération utilisée pour rejoindre la position idéale définie par les trois règles. Ces principes ont été utilisés pour la première fois dans Le retour de Batman en 1992, pour générer des vols de chauves-souris.
Le modèle peut être amélioré en limitant le voisinage à un secteur de cercle, correspondant à la vision de l’oiseau, à la considération d’obstacles que l’oiseau évitera et également aux prédateurs éventuels.
Pour chercher leur nourriture et la rapporter dans leur fourmilière, les fourmis suivent le chemin le plus court. Quand on les voit se déplacer l’une derrière l’autre en longues files indiennes, elles semblent obéir aux ordres donnés par leur reine cachée au fond du nid.
L’intelligence des fourmis
En fait, elles procèdent par essais et erreurs au niveau collectif de la fourmilière. La méthode pour cela est très simple. Une fourmi découvrant de la nourriture retourne à la fourmilière en déposant sur le sol une substance volatile appelée phéromone. Cette fourmi est rarement seule à faire cette découverte. Celle qui a trouvé le chemin le plus court rentrera plus vite et sera donc plus vite imitée. Le nombre de traces dans sa direction sera bientôt prépondérant et des colonnes de fourmis emprunteront sa trace tandis que les autres seront délaissées. C’est de cette façon que les fourmis déterminent le plus court chemin entre leur fourmilière et la nourriture. Elles se trompent rarement.
Fourmis virtuelles en quête d’un plus court chemin.
Quand l’intelligence artificielle s’inspire de celle de la fourmillière
Des informaticiens ont tenté, et réussi, de simuler le comportement des fourmis. Ils essayent aujourd’hui de résoudre des problèmes de plus courte distance au moyen de fourmis virtuelles. Ainsi, après avoir essayé d’imiter le comportement humain, l’intelligence artificielle suit aujourd’hui la voie des fourmis. On parle d’intelligence distribuée ou d’intelligence en essaim.
Les plantes ont un rapport étonnant avec les mathématiques, hasard ou nécessité ? Je vous laisse juger.
Suite de Fibonacci
Léonard de Pise, dit Fibonacci, a créé sa suite comme un simple exercice d’arithmétique :
Un homme met un couple de lapins dans un lieu isolé de tous les côtés par un mur. Combien de couples obtient-on en un an si chaque couple engendre tous les mois un nouveau couple à compter du troisième mois de son existence ?
Le calcul est simple, la suite donne : 1, 1, 2, 3, 5, 8, 13, 21, 34, 55, 89, etc. Chaque nombre est la somme des deux qui le précèdent.Cette règle a fasciné au-delà de l’exercice. De plus, on la retrouve souvent dans la nature. En voici quelques exemples.
Cette plante de la toundra groenlandaise présente deux formes géométriques simples : un cercle et une droite. Le cercle est naturel. Il correspond au développement de la plante dans toutes les directions à partir d’une graine, mais pourquoi a-t-elle dépéri d’un seul côté d’une droite ?
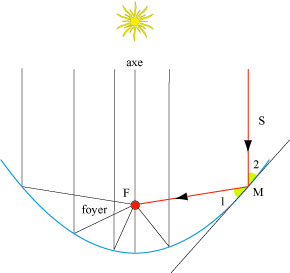
Les paraboles sont utilisées dans l’Himalaya pour faire bouillir de l’eau. Pour cela, il suffit de diriger son axe vers le soleil. Ses rayons sont alors réfléchis vers le foyer où on a placé une casserole.
La parabole et son foyer
Si le soleil est dans l’axe de la parabole, ses rayons réfléchis passent tous par le foyer.
Selon la légende, Archimède aurait utilisé ce procédé pour incendier les voiles des navires romains lors du siège de Syracuse en 212 avant Jésus-Christ. Nous pouvons douter de la réalité de cette anecdote, car le moindre mouvement des bateaux suffit pour placer leurs voiles loin du foyer. Les servants du miroir parabolique auraient bien du mal à les suivre. Il est plus facile de chauffer une bouilloire immobile que la voile d’un navire en mouvement !
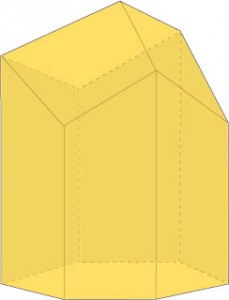
Les abeilles seraient-elles mathématiciennes ? Sans doute non mais elles sont étonnantes. Le gâteau de cire qu’elles construisent pour y déposer leur miel est formé par deux couches d’alvéoles opposées par leur fond. Dès l’antiquité, on avait remarqué que les alvéoles ressemblaient à des prismes droits à base hexagonale régulière. Ce n’est qu’au XVIIIe siècle que l’on remarqua que le fond était l’assemblage de trois losanges identiques appartenant chacun à deux alvéoles opposées.
Les alvéoles des abeilles sont des prismes de base hexagonale terminés par trois losanges inclinés, un peu comme un crayon taillé.
Une mesure, une hypothèse …
En 1712, Giacomo FilippoMaraldi (1665 – 1729), un astronome de l’observatoire de Paris, mesura l’angle des losanges et trouva : 109 degrés et 28 minutes. En 1739, René-Antoine Réaumur ( 1683 – 1757) soupçonna les abeilles de construire le fond de façon à utiliser le minimum de cire possible.
Et un calcul
Samuel König
Sans lui donner l’origine de son problème, il demanda de le résoudre à Samuel König (1712 – 1757), le mathématicien allemand connu pour avoir enseigné les mathématiques à la marquise Émilie du Châtelet (1706 – 1749), traductrice de Newton en français. König traita le problème par le calcul différentiel et, en utilisant une table de logarithmes, il en déduisit la valeur de 109 degrés et 26 minutes. L’erreur des abeilles était négligeable. On s’émerveilla de cette précision.
Un naufrage
À l’époque, les marins utilisaient la même table que König pour leurs calculs. Malheureusement, il fallut un naufrage quelques années plus tard pour que l’on y découvre quelques erreurs. En 1743, Colin Mac Laurin (1698 – 1746) corrigea la valeur trouvée par König : il s’agissait bien de 109 degrés et 28 minutes. La table de logarithmes avait tort et les abeilles, raison !
Comment comprendre le monde moderne sans culture mathématique ? Accéder à celle-ci n’exige cependant pas d’apprendre à résoudre la moindre équation.