Les coquilles des escargots sont des spirales qui peuvent croître de manière dextre ou senestre. En fait, ils sont presque tous dextres. Seuls un sur dix mille est senestre dans l’espèce des petits gris mais il existe des espèces où c’est le contraire.
Un petit gris. Si on place sa tête à gauche, sa coquille s’enroule dans le sens trigonométrique.
Les tire-bouchons usuels, c’est-à-dire pour droitiers, sont dextres, les tire-bouchons pour gauchers sont senestres.
Un tire-bouchon dextre.
Les queues de cochons
De même la queue en tire-bouchon des cochons peut être dextre ou senestre. Dans ce cas, il se trouve qu’il y a autant de cochons dextres que de cochons senestres. Le sexe des cochons mâles a la même propriété. Ces différences entre dextre et senestre se retrouvent au niveau des molécules, ce qui a parfois des conséquences sur leurs propriétés.
Le pangolin en boule
Pour échapper à ses prédateurs, le pangolin (oui, le mammifère impliqué dans l’origine du Covid-19) se roule en boule, ce qui le rend vulnérable aux braconniers.
Pangolin en boule
D’où la question : existe-t-il deux sortes de pangolins selon l’orientation de la boule ?
Dans leurs calculs, les statisticiens utilisent la loi des grands nombres. La française des jeux n’opère pas autrement pour gagner de l’argent ! Le hasard n’intervient que pour les joueurs, pas pour elle ! Les compagnies d’assurance agissent de même. Si elles assurent cent mille voitures, elles savent d’avance combien auront d’accidents et quel en sera le coût. La prime d’assurance est calculée en fonction de ce risque qui n’en est plus un dès que l’on applique la loi des grands nombres ! Si 5% des automobilistes ont un accident chaque année, vous ne pouvez prévoir si vous en aurez un. En revanche, votre compagnie d’assurance sait que, sur ses cent mille assurés, cinq mille auront un accident.
La loi des petits nombres
Les particuliers ne raisonnent pas ainsi. Si un événement malheureux mais peu probable se produit deux fois de suite à une année d’intervalle, ils se diront que jamais deux sans trois et prévoiront un troisième pour l’année suivante. A l’inverse, plusieurs années sans accident leur feront croire que plus rien ne peut leur arriver. Autrement dit, ils utilisent une loi des petits nombres et non la loi des grands nombres. Bien entendu, il ne s’agit pas de mathématique mais de psychologie !
Une question de psychologie
Pour un mathématicien, cette loi des petits nombres peut passer pour un canular. C’est pourtant de manière tout à fait scientifique et en utilisant correctement la loi des grands nombres que Daniel Kahneman l’a mise en évidence. Plus précisément, il a étudié expérimentalement le comportement moyen des américains devant l’assurance ! Il apparaît que plusieurs années sans accident pousse la moyenne des américains à résilier ses contrats d’assurance ! Pour cette étude, ce professeur de psychologie à Princeton a obtenu le Prix Nobel d’économie en 2002.
Il semblerait que certains états appliquent cette loi des petits nombres et suppriment des équipements de précaution, comme des masques de protection, quand ils se sont révélés inutiles plusieurs années de suite. D’autres, dans l’affolement, feront des tests de médicaments sur des petits nombres pour en déduire avoir trouvé le traitement miracle.
Le jeu de la vie, inventé en 1970 par John Conway, n’est pas vraiment un jeu. Ce terme est cependant moins rébarbatif que celui d’automate cellulaire, qui est pourtant plus exact. Il trouve ses origines dans des travaux conduits par John von Neumann dans les années 1940. Nous garderons la métaphore du jeu pour en parler, même si certains trouveront le terme mal adapté quand il s’agit de maladies potentiellement mortelles. L’essentiel est d’aider la compréhension. Voyons quelles en sont les règles.
Les règles du jeu de la vie
Pour jouer, prenez un damier et des pions. Les cases sont considérées comme des cellules ; elles peuvent être mortes ou vivantes. On utilise les pions pour matérialiser les cellules vivantes. Au début du jeu, on place des pions sur n’importe quelle case. On joue ensuite par étapes selon les règles suivantes :
— une cellule morte entourée de trois cellules vivantes ressuscite, sinon elle reste morte ;
— une cellule vivante reste en vie si elle a deux ou trois voisines vivantes, sinon elle meurt.
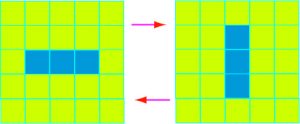
Bien que l’évolution du jeu soit complètement déterminée par la disposition initiale des cellules, on n’en assiste pas moins à quelques situations qui peuvent paraître surprenantes. Ainsi, en alignant tout simplement trois cellules vivantes les unes à côté des autres, on obtient une situation où les trois cellules se reproduisent, alignées horizontalement puis verticalement et ainsi de suite.
Lorsque trois cellules vivantes sont contiguës, on assiste à une oscillation entre trois cellules en ligne et trois en colonne.
Le jeu des épidémies
Ce jeu est loin d’être un simple amusement : il s’agit d’un exemple de ce que l’on nomme « automate cellulaire », particulièrement utile pour modéliser les processus d’expansion des épidémies comme des épizooties. En préalable à ce type d’application, il est nécessaire d’étendre le damier à l’infini. Au départ, toutes les cellules sont saines. On place une cellule infectée puis on « joue » avec la règle probabiliste suivante :
— les cellules voisines de la cellule infectée sont infectées au coup suivant avec la probabilité p ;
— la cellule meurt ou est immunisée le coup suivant.
Comment les cellules infectées (en rouge) se multiplient-elles au détriment des cellules saines (en vert) ? Dans cet exemple, la probabilité qu’une cellule voisine d’une cellule infectée soit infectée à son tour est de 25 %. Les cellules mortes ou immunisées sont représentées en bleu.
La question qui intéresse autant les épidémiologistes que le grand public est donc : « Pour quelles valeurs de p, la maladie se propage-t-elle au monde entier ? »
Un modèle probabiliste
Le modèle est ici « probabiliste », et donc on ne peut prédire à l’avance ce qui va se produire dans un cas particulier. Pour avoir une idée rapide de l’évolution moyenne du système, le mieux est de procéder à une simulation. Pour cela, on « joue » selon les règles énoncées ci-dessus en utilisant un générateur de nombres pseudo-aléatoires et on comptabilise le nombre de cellules infectées. En jouant cent fois de suite et en faisant la moyenne des résultats, on obtient une mesure de l’expansion moyenne de l’épidémie.
Taux critique
En dessous d’un certain taux de contamination p, l’épidémie ne s’étend pas. En revanche, au dessus de ce taux, elle envahit le monde entier. Dans le cadre de notre modèle simplifié, le taux critique se situe entre 30 % et 40 %. Une maladie ne devient épidémique que si ce taux est dépassé. Comment ce modèle peut-il être adapté pour bien modéliser différents types d’épidémies ou d’épizooties ? Tout d’abord, on peut modifier le voisinage de chaque cellule, composé ici de huit cellules — les spécialistes parlent de voisinage de Moore, du nom d’Edward Moore, l’un des fondateurs de la théorie des automates. On utilise souvent un voisinage plus simple, dit de von Neumann, constitué des quatre cellules partageant un côté avec la cellule considérée. Avec ce nouveau modèle, le taux critique pour lequel une maladie devient épidémique se situe aux alentours de 60 %. On peut également améliorer le modèle en tenant compte du temps pendant lequel une cellule infectée est contagieuse puis du taux de mortalité et d’immunité ainsi que du temps d’immunité. On arrive ainsi à retrouver la façon dont se sont propagées des épidémies comme la peste dans l’Europe médiévale. Une première vague a tué le tiers de la population en se propageant à partir d’un épicentre situé dans un port, suivie de plusieurs répliques plus faibles, toutes partant du même point. Ces répliques correspondent à la fin de certaines immunités.
La confrontation avec les données épidémiologiques a permis de montrer que ce type de modèles a une certaine pertinence pour toutes les maladies qui se propagent par contact direct : grippe, tuberculose, coronavirus ou même sida. En revanche, il ne fonctionne plus lorsque la maladie se propage via un agent infectieux, comme dans le cas du paludisme ou du chikungunya.
Géométrie des contagions
Comment considérer maintenant la notion de « cellule voisine » dès que l’on évoque les réseaux de transports aériens, maritimes ou terrestres ? Dans le cas d’une épidémie de grippe humaine, l’aéroport de Paris est voisin de celui de Hong-Kong. Dans le cas d’une épizootie de grippe aviaire, deux élevages fréquentant le même marché aux bestiaux sont voisins. On doit de plus tenir compte des migrations naturelles des oiseaux sauvages. Dans tous ces cas, on retrouve la notion de réseaux.
En modifiant le modèle du jeu, on peut passer du cas où chaque cellule représente un individu à celui où elle représente un domaine où les individus sont en relation constante : un élevage de volaille dans le cas de la grippe aviaire, une ville dans le cas de la tuberculose, du sida ou de la grippe humaine. Ces domaines sont reliés entre eux pour former un réseau. Dans chaque cellule, la modélisation suit une autre logique, celle du modèle « SIR » dû à William Kermack et Anderson Mac Kendrick en 1927 (voir l’article correspondant sur ce blog). Ce modèle compartimente la population en trois classes : S, la classe des individus susceptibles d’attraper la maladie, I, celle de ceux qui en sont infectés (et contagieuses) et R, ceux qui en sont guéris (et immunisés) ou décédés.
Seuil de propagation
On considère l’évolution de ces trois classes dans le temps en fonction de deux taux mesurables expérimentalement. Le premier (a) est le taux de contagion de la maladie pour un infecté, c’est-à-dire la probabilité pour qu’un individu susceptible attrape la maladie après contact avec un individu infecté. Le second taux (b) mesure le passage de l’état I à l’état R.
Après un laps de temps t, on compte aI S t infectés supplémentaires et R augmente de bI t. La variation du nombre d’infectés est donc égale à aS – b multiplié par I t. La condition pour que la maladie se propage (et donc donne lieu à une épidémie) est que le nombre de malades infectés augmente, c’est-à-dire que : aS – b > 0. Le quotient b / a a donc valeur de seuil. Si le nombre de sujets susceptibles est strictement inférieur à ce seuil, la maladie ne s’étend pas. Sinon, elle donne lieu à une épidémie (ou à une épizootie).
D’une façon qui peut paraître paradoxale, l’apparition d’une épidémie ne dépend donc pas du nombre de personnes infectées mais du nombre de personnes susceptibles d’attraper la maladie ! Cette remarque justifie à elle seule les politiques de vaccination, même avec un vaccin peu efficace.
L’ADN (ou acide désoxyribonucléique) est le support de l’hérédité. Cette molécule, présente dans chaque cellule, prend la forme d’une double hélice, qui s’enroule sur elle-même, formant ainsi un nœud.
Molécule d’ADN formant un nœud. Sa réplication demande de le dénouer. Image réalisée au moyen d’un microscope électronique.
Duplication des molécules
La duplication des informations contenues dans une molécule d’ADN se fait au moyen d’enzymes. Pour « voir » le processus, imaginez une longue fermeture éclair qu’on ouvre avant de la séparer en deux. Cela n’est possible que si le nœud peut être dénoué. Certains virus attaquent les molécules d’ADN en les coupant et en les recollant de sorte qu’ils soient impossibles à dénouer. Le type de nœud obtenu après l’attaque virale est caractéristique de chaque virus. La signature de ces virus est de nature topologique !
Par ailleurs, cette question du dénouement est au cœur de la théorie mathématique des nœuds. Certains sont faciles à dénouer, d’autres bien plus compliqués, voire impossible (voir la figure ci-dessous). À l’envers de celle des virus, la seule méthode est celle qu’Alexandre le Grand employa pour dénouer le nœud gordien : couper la corde !
Deux nœuds. Pour défaire le vert, il suffit de faire glisser la boucle de gauche. Le second requière la méthode d’Alexandre et des virus, non autorisée en théorie des nœuds.
Nœuds et mathématiques
Mathématiquement, les nœuds sont des courbes fermées de l’espace de dimension trois, que l’on représente souvent comme une courbe plane. Elle a alors des points doubles, où il faut distinguer la branche « au-dessus » de celle « en-dessous ». Si en essayant de démêler un nœud, on passe à un autre, les deux nœuds sont dits équivalents. La théorie des nœuds consiste donc à étudier si un nœud est équivalent à une courbe non nouée, comme le cercle, et plus généralement si deux nœuds sont équivalents. Pour étudier ce type de problème, on essaye d’introduire des invariants, c’est-à-dire des objets mathématiques invariants quand on passe d’un nœud à un nœud équivalent. Henri Poincaré (1854 – 1912) en a trouvé un particulièrement subtil, que l’on appelle le groupe du nœud, malheureusement son étude est délicate.
Stephen Smale (né en 1930), William Thurston (1946 – 2012) et Mikhaïl Gromov (né en 1943) réunis lors de la conférence Clay sur la résolution de la conjecture de Poincaré, en 2010.
William Thurston a découvert une réalisation concrète de ce groupe, liée à la géométrie des espaces de dimension trois, ce qui lui a valu la médaille Field en 1982, et explique son implication en biologie ainsi que celles de Stephen Smale ou de Mikhail Gromov, spécialistes de ce domaine, souvent présenté très loin de toute application.
Jean-Henri Fabre est connu pour son observation des insectes. Excellent vulgarisateur, il est de ceux qui savent communiquer leurs passions. Les mathématiques en font partie.
Jean-Henri Fabre
Jean-Henri Fabre (1823 – 1915)
Bien que titulaire d’une licence de mathématiques, d’un doctorat en sciences naturelles et de plusieurs autres diplômes, Jean-Henri Fabre est un autodidacte comme il le rappelle lui-même :
Apprendre sous la direction d’un maître m’a été refusé. J’aurais tort de m’en plaindre. L’étude solitaire a sa valeur ; elle ne vous coule pas dans un moule officiel, elle vous laisse votre pleine originalité. Le fruit sauvage, s’il arrive à maturité, a une autre saveur que le produit de serre chaude ; il laisse aux lèvres qui savent l’apprécier un mélange d’amertume et de douceur dont le mérite s’accroît par le contraste.
Son côté autodidacte le rend attachant pour certains et agaçant pour d’autres. Quelques modernes lui reprochent aussi de ne pas avoir épousé les thèses de Darwin qui, en revanche, reconnaissait en lui un observateur incomparable. Il s’explique lui-même dans une de ses lettres à Darwin :
Vous vous étonnez de mon peu de goût pour les théories, si séduisantes qu’elles soient. Ce travers d’esprit, si c’en est un, tient un peu à mes longues études mathématiques qui m’ont habitué à ne reconnaître la vérité qu’à la lueur d’un irrésistible faisceau de lumière. Ne jurant par aucun maître, libre d’idées préconçues, peu enclin aux séductions des théories, je cherche avec passion la vérité, près à l’admettre quelle qu’elle soit et de quelque fait qu’elle vienne. Et comme moyen de recherche, je ne connais qu’une chose : l’expérience.
Par ailleurs, Darwin l’avait chargé d’expériences sur les insectes retournant à leurs nids. Les résultats se trouvent dans l’œuvre de Fabre. De façon générale, on trouvera la plupart des écrits de Fabre sur internet.
Fabre créationniste ?
Parmi les critiques modernes faites à Jean-Henri Fabre, certains le stigmatisent comme créationniste car il ne croyait pas à la théorie de Darwin, qu’il comparait à celle de la génération spontanée. À la défense de Fabre, il faut noter que la théorie originelle de Darwin n’était pas celle qui porte son nom aujourd’hui. Il s’agissait plutôt d’une transposition de la sélection des espèces domestiques, pratiquée depuis longtemps par les éleveurs, en une sélection naturelle sous l’effet de modifications du milieu. Autrement dit, il lui manquait l’explication qui viendra avec la découverte des gênes, par Gregor Mendel au début du XXe siècle. La théorie de l’évolution telle que nous la connaissons est postérieure de vingt ans à la mort de Fabre ! Comment peut-on lui reprocher de ne pas l’avoir reconnue ?
Mais l’essentiel n’est pas là, il est dans deux choses, sans parler de l’inélégance d’attaquer les morts, qui ne peuvent se défendre. Premièrement, il faut savoir ne pas se tromper d’adversaires. Les obscurantistes que sont les créationnistes ne sont pas les disciples de Jean-Henri Fabre. Ils sont dans des religions qui refusent la science, et malheureusement pas la violence. Deuxièmement, de Jean-Henri Fabre retenons plutôt l’exceptionnel talent de vulgarisateur. Pour finir sur une note poétique et liée à la question de l’évolution, voici l’un de ses commentaires sur la parade nuptiale des scorpions languedociens : La colombe a, dit-on, inventé le baiser. Je lui connais un précurseur : c’est le scorpion.
Parade amoureuse de scorpions languedociens. Dans un cas sur deux au moins, le mâle (à droite) finira dévoré par la femelle (à gauche), ce qui atténue l’impression romantique donnée par Fabre.
Jean-Henri Fabre a réussi à me faire regarder les scorpions autrement, c’est pourquoi je me souviens de cette remarque. Aujourd’hui, elle me fait m’interroger : selon la théorie créationniste, parler de précurseur d’une espèce a-t-il un sens ?
Descriptions et mathématiques chez Fabre
Dans ses souvenirs entomologiques, Jean-Henri Fabre dépeint les mœurs des insectes de manière vivante, en les ramenant souvent aux nôtres. Il décrit ainsi le carabe doré en nous emmenant d’abord visiter les abattoirs de Chicago pour comparer ensuite leur efficacité à celles des carabes dont on saisit mieux ainsi la férocité comme la voracité.
Le carabe doré, qui sera l’occasion d’une digression sur les mœurs humaines pour Jean-Henri Fabre.
Il conclut alors sur nos origines et notre avenir, avec l’abolition de l’esclavage et l’instruction des femmes, les deux voies du progrès moral selon lui. Cette façon de généraliser sera parfois critiquée plus tard, comme peu scientifique. Il est vrai que, par moments, Fabre concluait un peu vite. Par exemple, voici comment il décrit la toile d’une araignée, l’épeire :
Nous reconnaîtrons d’abord que les rayons sont équidistants ; ils forment de l’un à l’autre des angles sensiblement égaux […] les divers tours de spire […] avec les deux rayons qui les limitent, forment d’un côté un angle obtus et de l’autre un angle aigu […] d’un secteur à l’autre, ces mêmes angles, l’obtus comme l’aigu, ne changent pas de valeur, autant que peuvent en juger les scrupules du regard seul.
Fabre reconnaît alors une propriété caractéristique de la spirale logarithmique et en conclut que la toile de l’épeire épouse cette forme, ce qui est rapide surtout quand la mesure a été faite à l’œil. Ceci dit, cela n’enlève rien à la qualité de son travail, et il n’en reste pas moins que, du fait de sa construction, la toile prend une forme de spirale.
Scarabée sacré en train de confectionner une boule.
De même, c’est de manière très mathématique qu’il explique la forme de poire que le scarabée sacré donne à la bouse dans laquelle il dépose son œuf : une sphère pour minimiser la surface externe afin de réduire la dessiccation, qui rendrait la bouse immangeable pour la larve, coiffée d’une sorte de cylindre contenant l’œuf, qui se trouve ainsi dans un endroit plus aéré.
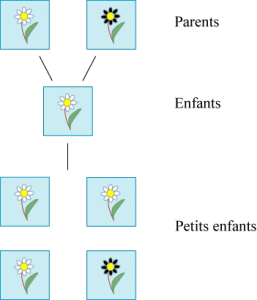
Gregor Mendel (1822 – 1884) est connu pour avoir posé les premières lois de la génétique. Elles sont de nature si mathématique que Godfrey Hardy, le grand mathématicien britannique du début XXe siècle, connu pour sa critique des mathématiques appliquées, les a prolongées. Imaginons qu’une fleur vienne en deux couleurs : blanche et noire, jamais grise ou autre et que ces deux variétés puissent s’hybrider, c’est-à-dire se mélanger. Imaginons que deux parents à fleurs blanches donnent toujours des enfants à fleurs blanches, alors que les parents à fleurs noires peuvent donner des blanches comme des noires.
Né Johann Mendel en 1822, Mendel prendra le prénom de Gregor à son entrée au monastère de Brunn (Tchéquie), en Autriche à l’époque. Il y trouva un milieu intellectuel stimulant et put y installer un jardin expérimental, où il fit ses recherches sur l’hybridation. En 1866, il devint supérieur de son couvent, ce qui mit fin à ses recherches en botanique. Il se consacra alors à l’administration du monastère ainsi qu’à des recherches en météorologie, pour lesquelles il fut reconnu par ses contemporains … davantage que pour ses apports à la génétique.
La mathématique de l’hybridation
Gregor Mendel a étudié ces lois de l’hybridation en pollinisant artificiellement des pois, qui se présentent sous deux formes facilement discernables. Nous ne décrirons pas ses expériences en détail. Son premier résultat est d’ordre statistique. En croisant une fleur noire et une fleur blanche, à la première génération, on obtient des fleurs blanches et, à la seconde, trois quarts de fleurs blanches et un quart de fleurs noires.
Deux premières générations d’un croisement blanche / noire.
Pour le mathématicien, une explication logique est de penser que le gène de la couleur des fleurs se divise en deux moitiés, ses deux allèles : blanc et noir. A priori, il existe donc quatre combinaisons possibles de ces deux allèles : blanc / blanc, blanc / noir, noir / blanc et noir / noir. Cette propriété est cachée car seuls les porteurs du gène blanc / blanc ont des fleurs blanches, tous les autres ont des fleurs noires. C’est pourquoi on parle de caractère dominant pour la couleur noire, et de caractère récessif pour la couleur blanche. Cette domination est cependant très relative car les combinaisons se faisant de façon équiprobable, à la seconde génération, nous trouvons une fois sur quatre la combinaison blanc / blanc, donc des fleurs blanches.
Cette théorie de Mendel ne fut pas comprise en son temps. Les biologistes pensaient que les caractères dominants devaient forcément augmenter dans la population, ce que les calculs précédents nient. Plus étrangement encore, on ne vit pas immédiatement le lien avec la théorie de l’évolution de Darwin, pourtant contemporaine de celle de Mendel.
Le principe de Hardy
À l’opposé de Mendel, qui était prêtre, Godfrey Hardy était un athée convaincu. Son athéisme comprenait cependant une étrange part d’autodérision, si on en croît l’anecdote suivante. La peur d’un naufrage lui fit écrire à un collègue pour lui annoncer qu’il avait démontré l’hypothèse de Riemann. Il aurait ensuite justifié son envoi en disant que Dieu, qu’il tenait pour son ennemi intime, n’allait pas le laisser mourir et laisser croire ainsi qu’un tel impie avait réussi à démontrer cette conjecture, encore ouverte de nos jours. Il est tout aussi étrange qu’un mathématicien pur aussi convaincu ait publié un article de biologie. Il le serait encore davantage si, un jour, il était plus connu pour son apport à la génétique que pour ses théorèmes mathématiques. Le moteur de recherche Google laisse penser que ce jour viendra puisque « théorème Hardy » donne 56 700 résultats alors que « principe Hardy » en donne 4 450 000. Godfrey Hardy y verrait sans doute une revanche de son ennemi.
Si les mathématiques appliquées ont pu un jour être vues comme « impures » par certains mathématiciens « purs », ce fut le cas de Godfrey Hardy. On s’étonnera alors de voir son nom mêlé à une question de biologie. C’est pourquoi il s’excusa presque de s’immiscer dans ce domaine. En 1908, au cours d’un dîner, on lui demanda s’il était possible de déterminer mathématiquement la proportion d’allèles dominants permettant l’évolution dans une population. Hardy étant un mathématicien pur, sa réponse réclama quelques hypothèses. Tout d’abord, la population devait être de grande taille, sans migration, estimée infinie, les individus s’y croiseraient aléatoirement mais les générations seraient séparées. Enfin, il n’y aurait ni mutation, ni sélection. Tout ceci assure la rigueur du raisonnement suivant.
Considérons un gène à deux allèles A et a possédant les fréquences p et q = 1 – p dans une certaine génération. Quelles sont les fréquences à la génération suivante ?
Pour le déterminer, comptons d’abord les fréquences des diverses combinaisons à la génération suivante : AA, Aa et aa. Il s’agit d’une question élémentaire de probabilité. Pour qu’un individu soit AA, il doit avoir reçu l’allèle A de ses deux parents, supposés aléatoires d’après l’hypothèse de Hardy. La fréquence de chacun étant égale à p, la probabilité est égale à p2. De même, celle de aa est q2. Pour Aa, deux cas sont possibles puisque cela peut provenir d’un A de la mère et d’un a du père, comme du contraire. On obtient donc 2 pq.
Si la population totale de cette nouvelle génération est égale à N, le nombre d’allèles y est égal à 2N. L’allèle A se trouve deux fois dans AA et une fois dans Aa, son nombre est donc égal à 2 p2 N + 2 pq N. Sa fréquence est ainsi égale à p2 + pq = p (p + q) = p puisque p + q = 1. Il en est de même de l’allèle a. Autrement dit, sous les hypothèses énoncées plus haut, la fréquence des allèles ne se modifie pas d’une génération à l’autre.
Ainsi, les relations de dominance entre allèles n’influent pas sur leurs fréquences. Autrement dit, l’évolution est impossible sous les hypothèses de Hardy … il faut tenir compte des mutations.
La même courbe se retrouve-t-elle dans les galaxies, certains mollusques et les toiles d’araignées ? Enquête sur la spirale logarithmique.
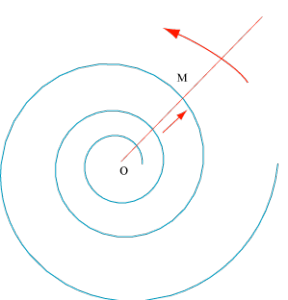
La spirale d’Archimède
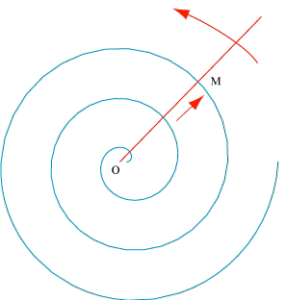
Imaginez ! Une droite tourne à vitesse angulaire constante autour d’un point O. Si, partant de O, un point M parcourt cette droite à vitesse constante, on obtient une spirale d’Archimède. On démontre facilement que les spires y sont régulièrement espacées.
Spirale d’Archimède. Elle est engendrée par un point mobile M partant d’un point O, à vitesse constante sur une droite tournant à vitesse angulaire constante autour de O.
La spirale logarithmique
Si, toujours partant de O, le point M parcourt la droite à une vitesse proportionnelle à la longueur OM, il dessine une autre courbe, appelée spirale logarithmique depuis Pierre Varignon (1654 – 1722) mais étudiée auparavant par René Descartes (1596 – 1650) avant d’être choisie par Jacques Bernoulli (1654 – 1705) pour orner sa tombe. Malheureusement, le sculpteur ignorait cette courbe et grava une spirale d’Archimède.
Spirale logarithmique. Elle est engendrée par un point mobile M partant d’un point O, à vitesse proportionnelle à OM sur une droite tournant à vitesse angulaire constante autour de O.
Au lieu d’être régulièrement espacées, les spires suivent une progression géométrique de raison constante. Autre propriété de la spirale : elle coupe le rayon OM suivant un angle constant.
Inscription sur la tombe de Jacques Bernoulli, avec la spirale en bas.Sur cet agrandissement, on voit que le sculpteur a gravé une spirale d’Archimède et non une spirale logarithmique. L’inscription latine “eadem mutata resurgo” signifie “déplacée, je réapparais à l’identique”.
Le développement du nautile
Le nautile est un mollusque marin dont la coquille est en forme de spirale. L’espace entre les spires étant triplé à chaque enroulement, elle évoque une spirale logarithmique. Pour examiner si cette forme est fortuite ou non, il est nécessaire d’en comprendre la provenance.
Coupe d’un nautile faisant apparaître une forme de spirale logarithmique.
La coquille du nautile est divisée en chambres closes, l’animal n’occupant que la dernière. Les autres sont remplies d’un mélange de liquide et de gaz, toutes communiquent entre elles au moyen d’un siphon.
Nautile vivant. L’animal n’occupe que la dernière chambre. Il se déplace d’avant en arrière en expulsant de l’eau du côté de sa bouche.
Ces chambres correspondent à l’évolution progressive du mollusque. Quand il grossit, ne pouvant agrandir la chambre où il se trouve, il en crée une autre dans son prolongement, un peu plus grosse mais semblable.
Pour montrer que cette idée mène effectivement à une spirale logarithmique, prenons comme modèle de la coquille une suite de triangles rectangles d’angle au sommet constant égal à 30°. Le rapport entre un triangle et son suivant est de 115 % (l’inverse du cosinus de 30° soit 2 divisé par racine de 3 pour être précis), ce qui correspond bien à une spirale logarithmique. L’idée correspond à un accroissement progressif de la taille de l’animal. Il n’est pas besoin d’imaginer de plans compliqués inscrits dans les gènes du nautile pour cela, juste une façon de croître.
Suite de triangles rectangles formant une (approximation de) spirale logarithmique.
La spirale logarithmique se retrouve pour les mêmes raisons dans d’autres animaux, comme la planorbe, un escargot marin très utilisé dans les aquariums car il se nourrit d’algues et de plantes à la limite du pourrissement.
Une coquille de planorbe en forme de spirale logarithmique.
Les toiles d’araignées
La toile d’araignée est avant tout un piège destiné à attraper des insectes. Certaines espèces tissent des toiles où il est bien difficile de reconnaître la moindre régularité.
Il n’est pas facile de reconnaître la moindre courbe mathématique dans cette toile d’araignée. En revanche, sans le soleil en contre jour, il est difficile de la détecter.
Les espèces les plus communes en France, les épeires, fabriquent cependant des toiles en forme de spirales. Après avoir bâti un cadre entre quelques branches, l’araignée tisse un réseau régulier de segments rectilignes partant tous d’un même point. Un fois ce travail fini, elle forme une spirale en les reliant. Le célèbre entomologiste Jean-Henri Fabre (1823 – 1915) a voulu y reconnaître une spirale logarithmique, tout en remarquant que l’action de la pesanteur transformait chaque segment en chaînette, la forme que prend naturellement un fil pesant comme les câbles électriques ou les chaînes que l’on porte autour du cou.
Cette toile d’épeire laisse plus penser à une spirale d’Archimède qu’à une spirale logarithmique. On y remarque également les segments transformés en chaînette sous l’effet de la pesanteur.
Le passé a connu des épidémies terribles comme les pestes du Moyen-Âge. Avant qu’on comprenne leur mode de transmission, les hommes étaient incapables de s’en prémunir. Quand on le comprit, on put opérer des mises en quarantaine. Enfin, la solution vint avec les vaccinations, qui permettent de réduire le nombre de gens susceptibles de contacter une maladie. Cela suffit pour éviter une épidémie, comme l’explique le modèle SIR. En l’absence de vaccin, on revient aux quarantaines mais avec un peu plus de souplesse.
Le modèle SIR
William Kermack (1898 – 1970) et Anderson Mac Kendrick (1876 – 1943).
William Kermack et Anderson Mac Kendrick ont modélisé les épidémies en 1927. Leur modèle compartimente la population en trois classes : S, la classe des individus susceptibles d’attraper la maladie, I, celle de ceux qui en sont infectés (et contagieux) et R, ceux qui en sont revenus ou morts. Dans les deux cas, ces derniers sont immunisés et ne contamineront plus personne. Le modèle SIR considère l’évolution de ces trois classes dans le temps en fonction de deux taux mesurables expérimentalement. Le premier (a) est le taux de contagion de la maladie pour un infecté, c’est-à-dire la probabilité pour qu’un individu susceptible attrape la maladie après contact avec un individu infecté. Le second taux (b) mesure le passage de l’état I à l’état R. Après un laps de temps Δt, on compte aI S Δt infectés supplémentaires et R augmente de bI Δt. La variation du nombre d’infectés est donc égale à aS – b multiplié par I Δt. La condition pour que la maladie se propage (et donc donne lieu à une épidémie) est que le nombre de malades infectés augmente, c’est-à-dire que aS – b > 0. Le quotient b / a a donc valeur de seuil. Si le nombre de sujets susceptibles est inférieur à ce seuil, la maladie ne s’étend pas. Sinon, elle donne lieu à une épidémie (ou à une épizootie).
Un effet de seuil
D’une façon qui peut paraître paradoxale, l’apparition d’une épidémie ne dépend donc pas du nombre de personnes infectées, mais du nombre de personnes susceptibles d’attraper la maladie !
La vaccination permet de passer en dessous du seuil qui permet une épidémie.
Cette remarque justifie à elle seule les politiques de vaccination, même avec un vaccin peu efficace. Pour éviter une épidémie, c’est le nombre d’individus susceptibles d’attraper la maladie qu’il faut diminuer ! Ce nombre dépend de chaque maladie. Pour la rougeole, pour éviter l’épidémie, une couverture vaccinale de 95 % est nécessaire, ce qui n’est plus assuré en France du fait de campagnes obscurantistes antivaccins. La France a donc la honte d’exporter la rougeole dans des pays comme la Suisse ou le Costa Rica.
Calcul du seuil
Le calcul du seuil est très délicat. Pour cela l’usage est d’introduire un autre coefficient, appelé le taux de reproduction de base et noté R0 (R zéro) qui est le nombre de cas secondaires produits par un individu infectieux moyen au cours de sa période d’infectiosité, dans une population entièrement constituée de personnes susceptibles d’attraper la maladie. D’après la définition même, il y a épidémie si et seulement si R0 > 1. Un beau résultat … qui ne signifie rien si on ne sait pas évaluer ce R0 ! Ainsi, au début de l’épidémie de Covid-19, il était de 3,3 selon l’institut Pasteur et descendu à 0,5 après le confinement. Ces simples résultats montrent que la méthode ne vaut que pour un territoire uniforme et qu’une autre logique doit s’appliquer quand les cas sont rares. C’est d’ailleurs bien le cas puisqu’on essaye de confiner les foyers de contaminations isolés. En revanche, les modèles utilisés supposent une population homogène, ce qui n’est absolument pas le cas. Les personnes affaiblies par le grand âge ou des comorbidités risquent plus de contracter une forme grave de la maladie tandis que les jeunes sont souvent plus insouciants et, de ce fait, risquent davantage d’être contaminés mais avec des formes bénignes ou même asymptomatiques. Le modèle SIR est-il applicable dans le cas du Covid ? Sans doute non car il suppose un taux de contamination indépendant des conditions climatiques et homogène dans la population ce qui semble faux. Sans doute faudrait-il considérer la saisonnalité et des groupes d’âge, ou de la quantité de virus à laquelle on a été exposé, qui implique la charge virale. D’autre part, les données collectées pour calculer le R0 sont sujettes à caution. En particulier, les chiffres en provenance de Chine au début de l’épidémie laissaient prévoir une létalité très faible. Tout au plus, de 0,04 % ce qui n’est guère inquiétant. Elle fut en fait bien plus importante et très variable selon les pays, d’autant plus que les méthodes de collectes des décès différentes selon les pays biaisent les résultats. Pour conclure, les modèles sont importants d’un point de vue théorique mais peuvent être peu fiables en pratique. S’y fier aveuglément peut être dangereux, il importe de garder son bon sens et ne pas tuer une population atteinte sous prétexte de la sauver. La survie dépend aussi de l’économie ! De même, comment évaluer les traitements quand 98 % des personnes infectées guérissent sans le moindre traitement ? Le problème se pose également pour l’évaluation d’un vaccin.
Les mesures de confinement
En l’absence de vaccin, il reste qu’on peut essayer de diminuer le taux a ce qui augmente le seuil b / a. Pour cela, on peut éviter une quarantaine stricte et se contenter de réduire les interactions sociales. Dans le cas du Covid19, il semble que limiter les interactions sociales à 5 par jour suffit.
Le théorème du moustique
Le modèle SIR justifie également le théorème du moustique découvert par Ronald Ross (1857 – 1932) en 1911, et selon lequel il n’est pas besoin d’éliminer tous les moustiques pour éradiquer le paludisme, il suffit d’en faire passer la population sous un certain seuil. Cette découverte a précédé sa justification théorique au moyen du modèle SIR. Auparavant, le lien entre les marécages et le paludisme était connu, comme le montrent les tentatives de drainage des marais Pontins près de Rome de l’Antiquité jusqu’au XIXe siècle. De même, le paludisme a disparu de France, où il était autrefois endémique dans les régions humides comme la Sologne, le marais Poitevin et même autour de Port-Royal des Champs, au cours du XIXe siècle grâce au drainage et non à la consommation de Quinquina, un apéritif à base de Quinine, un préventif du paludisme, comme cela a été parfois affirmé !
Voir aussi : https://blogs.futura-sciences.com/lehning/2020/03/19/le-jeu-de-la-vie-et-celui-des-epidemies/
Comment comprendre le monde moderne sans culture mathématique ? Accéder à celle-ci n’exige cependant pas d’apprendre à résoudre la moindre équation.